THC (TETRAHYDROCANNABINOL) ACCUMULATION IN GLANDS OF CANNABIS (CANNABACEAE)
Paul G. Mahlberg and Eun Soo Kim, Department of Biology, Indiana University, Bloomington, IN USA; and Department of Biology, Konkuk University, Seoul, Korea
INTRODUCTION
THC (delta 9-tetrahydrocannabinol) is known to be present in flowering plants of Cannabis. However, its location in the plant and particularly in the cell remains less generally known. Our studies have been directed to determining where these compounds are localized at the whole plant and cellular level, with a long term objective to determine the organelle or membrane in the cell in their synthesis. It also should be possible to identify the gene responsible for synthesis of these compounds, THC in particular, and modulate this gene so as to develop strains of Cannabis with no THC, or no cannabinoids. Such strains would be intended for hemp agriculture.
The first phase of this study is to determine localization of THC in the plant. As part of this program we initiated an effort to accumulate a germplasm collection of various strains of world-wide distribution that are utilized in hemp cultivation in the classical sense, as well as seed oil strains and those with various levels of THC. We have utilized these strains for analyses of cannabinoids and their distribution; they also will serve as a germplasm source for future studies.
The purposes of this study are: a) to determine where cannabinoids are localized in the plant, and in which specific tissue, and b) to determine where within the cell or tissue cannabinoids are localized. Preliminary studies show it to be in the gland. Historically, it has been reported that a glandless mutant was detected at one time, but is now lost. If THC is in the gland, and glandless mutants can be produced, it should then be possible to reduce significantly the THC content of the plant. Such a plant, with its low THC content, would be a potentially important strain for the industrial hemp industry.
MATERIALS AND METHODS
For the localization studies, typically a higher THC strain (6%) was employed, although in some phases comparisons were made with a low THC or fiber strain, and with one possessing an intermediate level of THC.
Plants were grown from seeds in pots under greenhouse conditions. Flowering plants were developed by subjecting them to short-day (8 hr. daylength) by covering the bench with a black cloth. Plants were maintained in the vegetative condition by extending daylength to 16 or more hours by supplementing daylight with artificial lighting (Hammond and Mahlberg, 1973).
Data on morphological comparisons were obtained from populations of 10 or more plants maintained under a specific condition. Since plants from the same seed sources were grown many times under these conditions we had repeated opportunities to examine the morphological features during our studies. Features examined included seedling development, leaf morphology, flowering character of both pistillate (female) and staminate (male) plants, individual flower characters, glands and hairs on leaves and floral bracts at all stages in development (Hammond and Mahlberg, 1977, 1978; Kim and Mahlberg, 1991; Mahlberg, et al., 1984; Mahlberg and Kim, 1991, 1992).
Phenol contents of glands and other tissues were examined by thin layer chromatography (TLC), gas liquid chromatography-mass spectroscopy (GC-MS) and nuclear magnetic resonance spectrometry (NMR), and terpenes were analyzed by GC-MS (Hammond and Mahlberg, 1992, 1990, unpublished).
For gas liquid chromatography (GC) weighed samples of leaf material at similar stages in development from different plants, weighed samples of bract or other floral materials also at similar stages in development, isolated contents of whole glands or only the secretory contents of the non-cellular cavity of the gland were extracted with ether or chloroform and evaporated to dryness. Samples were redissolved in a given quantity of absolute alcohol containing the internal standard, androstenedione (1 mg/ml), and injected onto the GC column. Analyses were performed on a Hewlett-Packard 5710A chromatograph equipped with a hydrogen flame-ionization detector and programmed from 200-240 C (2 C/min). Nitrogen (20 ml/min) was used as the carrier gas. Injection port and detector temperatures were 250 and 300 C, respectively. Glass columns were packed with 3% OV-1 Supelcoport (80/100 mesh). Quantitative data were determined with a Hewlett-Packard integrator (3380A), and samples were compared with cannabinoid standards obtained from the National Institute on Drug Abuse (Hemphill, Tuner and Mahlberg, 1980; Turner and Mahlberg, 1984; Turner, Hemphill and Mahlberg, 1980).
Whole gland heads were removed individually from leaf or bract surfaces under a stereomicroscope with a tungsten microneedle and placed in vials containing chloroform to extract cannabinoids. The needle was rinsed in chloroform after collecting each gland to avoid cross contamination between collected glands. Samples of twenty and 100 glands were collected for different sampling periods and analyzed by GC as described (Turner, Hemphill and Mahlberg, 1978).
Contents of the secretory cavity were obtained by glass microcapillary micromanipulation. Bracts containing numerous glands were mounted on a special slide holder under a microscope lens. Microcapillary tubes, made with a Knopf micropipet puller and ground with diamond paste to a hypodermic tip, were mounted in a modified Leitz manipulator system under a microscope lens to probe into the secretory cavity without touching or damaging the disc cells. Microcapillary tubes with secretory cavity contents were placed in small acetone-washed test tubes containing solvent to dissolve contents, and subsequently prepared for GC analyses (Lanyon, Turner and Mahlberg, 1981; Turner, Hemphill and Mahlberg, 1978).
For scanning electron microscopy, bracts or other tissue were fixed in a buffered glutaraldehyde solution, rinsed in buffer, dehydrated through an ethanol series to acetone and critically point-dried. Samples then were mounted on specimen holders and examined at 20 kV with a Cambridge 250 scanning electron microscope (SEM) (Hammond and Mahlberg, 1973).
For transmission electron microscopy (TEM), bracts with glands were prepared by high pressure cryofixation-cryosubstitution for the THC antibody procedure. These tissues were fixed under cryo-conditions in glutaraldehyde alone or in glutaraldehyde and osmium tetroxide, embedded in resin, sectioned and treated with the antibody probe, stained and examined under a TEM at 60 kV. The monoclonal antibody for THC, obtained from Roche Diagnostic Systems, was prepared in mouse ascites and is normally used to screen humans for drug abuse. We developed it as an electron microscopic probe. Antibody-treated sections were visualized for THC with protein-A bound gold particles (20-nm, for Jannsen, Belgium). Controls included sections treated with antibody alone, or treated with protein-A-gold alone (Kim and Mahlberg, 1997a). For TEM developmental studies, bracts at different stages in development were fixed in glutaraldehyde, post-fixed in osmium tetroxide and embedded in resin. Thin sections were stained with lead citrate and examined at 60 kV (Kim and Mahlberg, 1991, 1995, 1997b; Mahlberg and Kim, 1991, 1992).
RESULTS AND DISCUSSION
Cannabinoids and THC
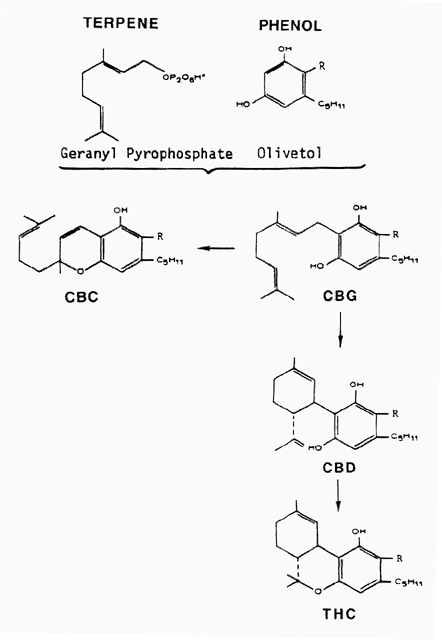
In this study we will examine two types of the several glands on Cannabis, the capitate sessile and capitate stalked forms. In addition, there are hairs on the surface of the plant. In our commentary we will refer to cannabinoids as the broad class of specialized compounds synthesized by Cannabis. Cannabinoids are dimers in that they are formed by condensation of terpene and phenol precursors (c).
Diagram 1 (Above). Cannabinoid pathway. Cannabinoids represent a dimer consisting of a terpene and a phenol component. Cannabigerol (CBG) is the first component of the pathway. It undergoes chemical change to form either cannabichromene (CBC), or cannabidiol (CBD). Delta 9-tetrahydrocannabinol (THC) is derived from CBD.
Cannabinoids include these more abundant forms:
THC, delta 9-tetrahydrocannabinol
CBD, cannabidiol
CBC, cannabichromene
CBG, cannabigerol
Another cannabinoid, cannabinol (CBN), is formed from THC and can be detected in some plant strains. Typically, THC, CBD, CBC and CBG occur together in different ratios in the various plant strains. In fiber strains CBD/CBC are in high concentrations and THC is at a low concentration; in drug strains, THC is high and CBD/CBC are low. An important area to be studied relates to the occurrence in the plant of potential precursor for cannabinoid formation and where synthesis occurs in the cell.
Distribution and types of glands
Glands cover entire surface of the above ground portion of both pistillate (female) and staminate (male) plants, but are most abundant on bracts of female plants (Fig. 1).
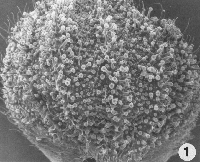
Figure 1. (Above) Stalked and sessile glands on underside of a bract. Hairs also are present on bract. x35.
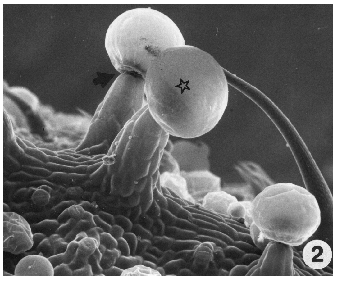
Figure 2. (Above). Stalked gland showing large head (star) and abscission zone at base of gland head (arrow). Sessile glands are evident in background along with a portion of a hair. X300.
Legend for all figures: C, cuticle; D, disc cell; P, plastid; S, secretory material; T, secretory cavity; V, vesicle; W, disc cell wall.
There are two types of glands active in cannabinoid secretion on female plants (Figs. 1, 2):
a. Capitate sessile - most common in that it occurs on stems, leaves and bracts.
b. Capitate stalked - develops only after flower formation, and occurs especially on the bracts subtending a flower and seed. Both types are present on the bract subtending the seed, but some factor(s) that stimulates flowering also stimulates development of the stalk related to this gland. Thus this gland has evolved from the sessile type.
An abscission zone develops at the base of the head where the stipe cells attach to the disc cells resulting in abscission of glands upon attaining maturity (Fig. 2, arrow).
Cannabinoid, including THC, composition of glands
a. Effect of position and season on contents of gland.
We examined both gland types for their cannabinoid composition. Whole glands (20 glands) were removed from bracts and leaves of the same plant to compare their contents. They were extracted for cannabinoids and analyzed (Table 1).
We also separated them as to glands over a vein as contrasted to non-vein areas. Results showed that stalked glands over a vein on the bract contained more THC content, approximately 20 times more, than the sessile glands over a leaf vein. Similarly, stalked glands over a non-vein area contained much more THC than sessile glands over the leaf non-vein area.
Surprisingly, even stalked glands on a bract varied for THC content between the vein and non-vein area. We also found that sessile glands varied for contents between vein and non-vein areas on the leaf.
This table shows two intervals during the year, October and December, during which we compared levels of THC in glands. Comparison of October with December analyses showed again that stalked glands contained considerably more THC than sessile glands. For both types, however, the vein glands now showed less than the non-vein. Interestingly, the level of THC can decrease to a non-detectable level in the glands (leaf vein glands).
In similar analyses of these glands on other strains we found the same pattern, but the cannabinoid levels in the stalked glands were not as high as for this strain. In this strain the sampling was repeated during the following March and, again, the stalked glands on the bract contained higher concentrations of THC than sessile glands on the leaf. Thus, a repeatable pattern appears to occur in the plant in which stalked glands usually contained greater quantities of THC than sessile glands.
b. Effect of gland age on cannabinoid content
We also examined cannabinoid content of stalked gland by age to measure the major cannabinoid components in both a fiber and drug strain (Table 2). Glands, viewed under a microscope, can be classified according to their secretory phases from the color of their contents. Glands most active in secretion (mature) are translucent in appearance, aged glands are yellow in appearance and senescent glands are brown in color. Mature glands possessed the highest content of their major cannabinoid in both the fiber and drug strains. Senescent glands possessed low levels of cannabinoids. The concentration of some components, as CBD in the drug strains, may be so low that is was not detectable in our analysis; similarly, for THC and CBN in the fiber strain. It is unknown where the cannabinoids go during the aging process, but we suggest that it is possible they volatilize into the atmosphere along with the terpenes in glands, as noted later in this report. Nevertheless, this phenomenon of altered content in glands during aging is one that should be studied to gain a more complete understanding of the secretory process of cannabinoids in the cell.
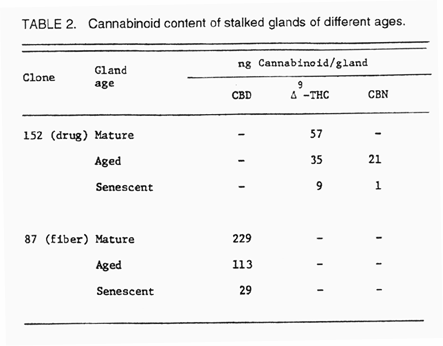
Table 2. (Above) Cannabinoid content of capitate-stalked glands of different ages.
c. Content of secretory cavity
The gland consists of disc cells with their cytoplasm and a non-cellular intrawall secretory cavity. We examined the contents of this secretory cavity specifically to determine if cannabinoids occur in it. With microcapillary pipets we removed contents from only the secretory cavity without damaging the disc cells. Data show cannabinoids to be abundant in this cavity (Table 3). Each gland contained an average of 61 nanograms of cannabinoids. This strain was a hemp form with a CBD content. In these analyses each sample included the contents of approximately 100 or more glands. Some samples represent bracts of different lengths, ranging from 4 to 9 mm, the latter being the mature size. Those bract samples of small size, 4 mm., showed a range of variation in cannabinoid content per gland, but some showed the highest content (samples 12 and 16) while others showed a low content (sample 7). Similarly, samples from larger bracts showed relatively high content (sample 6) as well as low content (samples 1 and 2). Some samples also showed different ratios for CBD to THC even when total cannabinoid content was high (compare samples 12 and 16).
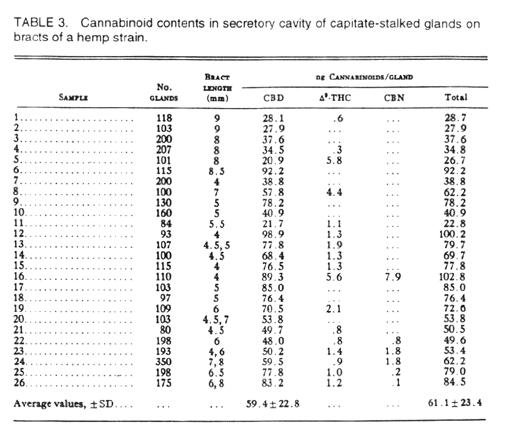
Table 3. Cannabinoid contents in secretory cavities of capitate-stalked glands of a hemp strain showing average cannabinoid content in a gland.
Although these difference perhaps should be expected in biological samples, it also emphasizes that the cellular component(s) which synthesize cannabinoids may not synthesize them in a constant ratio during development of an organ.
Summary:
a) Capitate stalked glands contained more THC (and total cannabinoids) than capitate sessile glands.
b) THC, and total cannabinoid, quantity in both gland types can vary during the year, and can decrease to very low levels.
c) Cannabinoids decrease with aging of glands.
d) Cannabinoids occur in the secretory cavity of the gland.
Location of cannabinoids in the gland
a. Development of the gland.
For both types of glands an epidermal cell enlarges and divides several times to form a tier of disc cells on a short stipe on the epidermis of the leaf or bract (Diagr. 2).
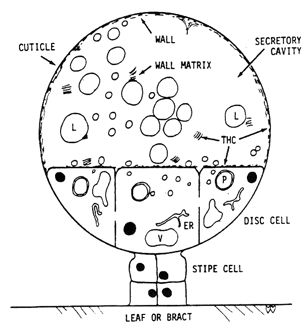
Diagram 2. (Above). Representation of mature secretory gland. Disc cells, attached to leaf or bract by stipe cells and basal cells (below stipe), release fibrillar wall matrix into secretory cavity where it contributes to thickening of subcuticular wall (wall) during enlargement of secretory cavity. Plastids (P) in disc cells produce secretions which accumulate outside plasma membrane, pass through cell wall as hyaline areas to form secretory vesicles (L) in secretory cavity. Vesicles in contact with subcuticular wall release contents to contribute to growth of cuticle during enlargement of secretory cavity. THC occurs in walls, fibrillar matrix and other contents surrounding the vesicles, but not in the vesicles; little THC is present in the disc cells. Nucleus = black; vacuole = V; vesicles = L; plastids = P; endoplasmic reticulum = ER.
The outer wall of the disc cells splits tangentially to initiate an intrawall cavity across the top of the entire gland surface. This cavity enlarges as secretions are accumulated in it (Fig. 3).
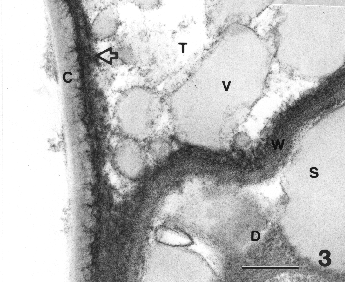
Figure 3. Section where secretory cavity joins disc cell showing cuticle and subcuticular wall, vesicles in secretory cavity, secretions in disc cell just below the wall of the disc cell separating it from the secretory cavity. Bar = 0.5 µm.
The outer portion of the wall remains associated with the cuticle to form the subcuticular wall; the inner portion remains associated with the disc cells. Both the cuticle and subcuticular wall increase in thickness as the secretory cavity enlarges and, therefore, precursors for their growth must be present in the secretory cavity. Secretions as vesicles are evident in the secretory cavity. Note the vesicles are similar in density (grayness) to gray secretions in the disc cell. The mechanism that controls the thickening of cuticle and subcuticular wall are as yet unknown.
b. Secretion role of disc cells
It is pertinent to examine the organization of the disc cells because all contents in this cavity must be derived from the disc cells. Cannabinoids, or their precursors, are secretions from these cells. Another major group of secreted compounds are the terpenes (monoterpenes and sesquiterpenes). Monoterpenes are the more abundant of the two. Terpenes compose the "essential oils"; they contribute to the odors of the plant, and are sticky in character, as evident when one touches the plant. Different combinations of terpenes in different strains contribute to odor differences among the strains. Cannabinoids, and THC, are odorless to most humans.
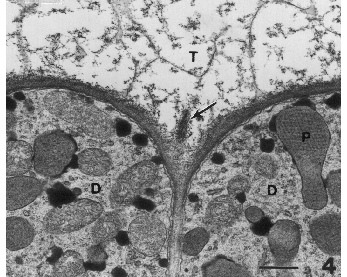
Figure 4. Portion of gland showing several disc cells each with numerous lipoplasts. Portion of secretory cavity is evident. Fibrillar matrix has separated from the wall (arrow). Bar = 0.5µm.
The tier of disc cells contains a typical cell complement including a large nucleus, plastids, mitochondria, endoplastic reticulum and abundant ribosomes, as well as vacuoles (Diagr. 2; Fig. 4). Plastids, however, represent the unique component of these cells. They are interpreted to be a source, perhaps the principal source, of secretions in the cavity. Plastids divide repeatedly and become very numerous in the disc cells. Plastids form a unusual central component, termed the reticulate body, derived from thylakoids (Figs. 4, 5). This body consists of thylakoids fused into a tubular array of light and dark areas in a hexagonal arrangement. This large body is somewhat like a prolamellar body that forms in chloroplasts when plants are grown in the dark because it has a lattice configuration similar to a prolamellar body. But it is unlike a prolamellar body in that it persists in both the light and the dark, and it does not contribute to formation of grana membranes as does the prolamellar body.
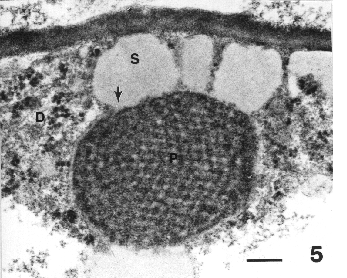
Figure 5. Lipoplast showing presence of secretion on its surface and region where secretion joins the reticulate body (arrow). Bar = 0.2 µm.
The reticulate body was associated with secretory activity. Enlarged quantities of secretions accumulated on the surface of the plastids, and were continuous with the light zone in the reticulate body (Fig. 5). The association of the secreted mass with the light zone on the surface of the plastid supported an interpretation that the reticulate body contributes to the synthesis of these secretions (Fig. 5, arrow). Such secretions were evident on nearly all plastids and may become so voluminous as to surround a plastid, as it appears in an electron micrograph. These secretions are more or less round in appearance in these sections, but undoubtedly spherical in three-dimensions; since they are oily in composition they form spherical masses in the aqueous medium of the cell. These secretions are interpreted to be terpenes; plastids in other plants are reported to produce terpenes.
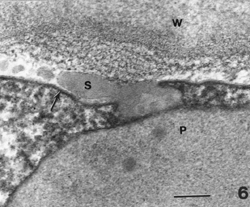
Figure 6. Lipoplast near plasma membrane (arrow). Portion of secretion in contact with and passing through the plasma membrane (arrow). It will accumulate between the membrane and cell wall. Bar = 0.1 µm.
The secretions on the surface of plastids can be observed in contact with the plasma membrane (Figs. 5, 6). Similarly, other quantities of secretions of similar density, but not in contact with a plastid in a thin section, were observed in contact with the plasma membrane (Figs. 3, 7).
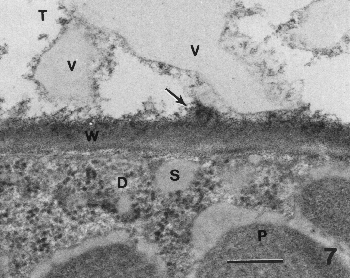
Figure 7. Secretions (small light areas in wall) emerging as a vesicle (V) in the secretory cavity (T). Fibrillar matrix (arrow) appears to be released from the wall into the secretory cavity. In disc cell, secretions are evident in the cytoplasm adjacent to the plasma membrane and on the surface of lipoplasts. Bar = 0.2 µm.
Secretions in the secretory cavity
Secretions somehow passed through the plasma membrane and accumulated in the space between the plasma membrane and cell wall (Fig. 6, arrow and S). Secretions subsequently passed through the cell wall, often appearing as small light areas in the wall (Figs. 3, 5, 6).
Secretions emerged into the secretory cavity as small accumulations on the wall surface facing the cavity (Figs. 3). Small gray vesicles, partly embedded in the wall, at W and to the lower left of the large vesicle (V) near the "corner" of the secretory cavity. They have the same gray density as secretions in disc cell (Fig. 6). It appeared that the material passing through the wall accumulated in enlarged vesicles on the wall surface facing the secretory cavity. As vesicles emerge from the cavity they became surrounded with a surface feature about Ω the thickness of a typical membrane (Figs. 3, 7).
Vesicles are released from the wall surface to aggregate in the secretory cavity (Fig. 8). These vesicles are transported to the subcuticular wall surface where some of them released their contents into the wall (Fig. 8, curved arrow). The contents also moved through the subcuticular wall to the cuticle where the contents aided thickening of the cuticle (Figs. 3, 8). The irregular contour of the inner surface of the cuticle with dark fiber-like extensions in the cuticle, is derived from the fusion of quantities of vesicular material along with its surrounding surface feature.

Figure 8. Outer sheath of secretory cavity consists of cuticle and subcuticular wall. Vesicles (light areas) are evident in the subcuticular wall (curved arrow). Vesicles and fibrillar matrix are evident in the cavity. Bar = 0.1 µm.
Fibrillar matrix is released from the surface of the wall into the secretory cavity. This matrix is evident at this wall as well as in the secretory cavity (Figs. 4 arrow, 7 arrow). The fibrillar matrix is transported to the subcuticular wall where it becomes incorporated thereby contributing to the thickening of this wall. The mechanisms controlling deposition of fibrillar matrix in the subcuticular wall, as well as the deposition of vesicular materials into the subcuticular wall and cuticle, remain to be studied. Since the sites of deposition are very distant from the origin of these materials, we speculate that the control mechanism somehow resides in the non-cellular secretory cavity.
Summary:
a) Plastids produce a major quantity of secretions that are released from their surface to pass through the plasma membrane and outer wall into the secretory cavity.
b) Secretions accumulate in the secretory cavity as secretory vesicles whose contents contribute to thickening of the cuticle.
c) Volatile components of secretory vesicles may volatilize into atmosphere and contribute to plant odor.
d) Fibrillar matrix and other material released from the wall surface surround the secretory vesicles with a surface feature of yet unknown composition.
Fibrillar matrix contributes to thickening of the subcuticular wall.
Localization of THC in the gland
Although cannabinoids were detected in the contents isolated from the secretory cavity, we do not yet know whether they occur in the disc cells. Further, we do not yet know where they are in the secretory cavity: in or around the vesicles, or elsewhere in the cavity.
For this phase of study fresh glands were frozen by high pressure cryofixation and then fixed (killed) by cryosubstitution to prevent movement of THC in the gland during the fixation process. We prepared a monoclonal antibody for THC as a probe by attaching gold particles to it so that it would be visible under the electron microscope. Then thin sections of glands were treated with the antibody probe; the antibody will attach to any THC in the tissue. Under the electron microscope we will see the electron dense gold particles, as dense black dots, where the antibody has attached to THC.
Upon examining the tissues we find that the THC, as indicated by the location of gold particles, is present in the cell wall facing the secretory cavity (W), and in the subcuticular wall (large arrowhead) under the cuticle (Fig. 9). It is also present in the fibrillar matrix being released into the cavity from the disc cell wall (open arrow). It was along the surface feature (small arrowhead) surrounding the large secretory vesicles in the cavity. It was also in the cuticle (arrow).
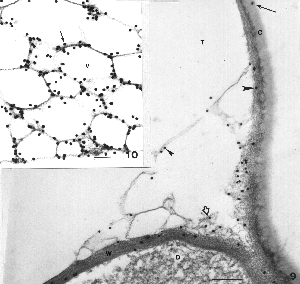
Figure 9. (Above) Sites of THC accumulation are evident as black dots representing the gold attached to the THC antibody. THC is present in the disc cell wall (W), subcuticular wall (large arrowhead), along the surface features around vesicles in the cavity (small arrowhead), in fibrillar matrix being released from the disc cell wall (open arrow) and in the cuticle (arrow). No particles were evident outside the gland. Bar = 0.2 µm. Figure 10. THC is abundant along the surface features (arrow) of the numerous vesicles in the secretory cavity, but absent from the content in the vesicles. Where the surface feature of a vesicle is sectioned in surface view, THC appears over the entire surface. Bar = 0.1 µm.
Deep within the cavity we note, again, that THC was associated with the surface feature around the numerous secretory vesicles, but it was not inside the vesicles (Fig. 10 arrow). Some vesicles were cut with their surface feature in planar view (slightly gray appearance), and gold grains are associated with the area of the surface feature. The contents of these vesicles (clear area), presumably, are the monoterpenes (lipoidal materials).
Surprisingly, it is not in the cytoplasm of the disc cells (Fig. 9). We detected it only along the plasma membrane and in the wall proper of the disc cell. Only an occasional grain was detected in the cytoplasm, either over a plastid, or mitochondrion or among ribosomes. If cannabinoids are synthesized in the cytoplasm of disc cells, there should be abundant gold particles at sites of THC synthesis and accumulation.
In other cells, as epidermal cells, THC was present in the wall, but in smaller quantities than in walls of the disc cell. Few gold particles were in the cytoplasm or vacuoles of other cells.
Controls included sections treated with antibody alone, or treated with protein A-gold alone. Controls showed no antibody in the disc cell wall, around vesicles, in the subcuticular wall or in cuticle. No antibody was detected outside of the cells.
Possible site of cannabinoid synthesis
Our studies contribute several pieces to the puzzle on where cannabinoids are synthesized, yet we lack definitive information on the precise site of their formation. Data from the antibody probe showing that they are not evident in the cytoplasm of the disc cells suggest that they may be formed at or outside the plasma membrane surface. A working model for continued study on their synthesis is embodied in diagram 3.
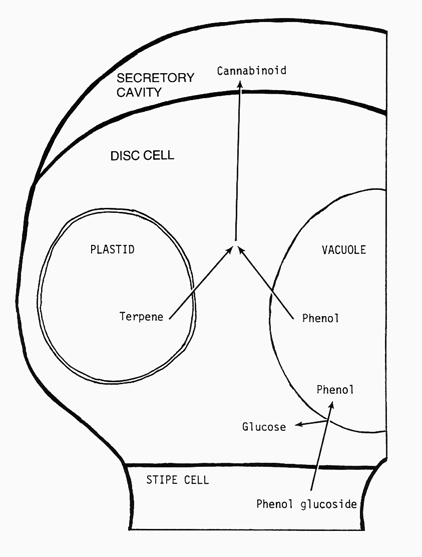
Diagram 3. (Left) Representation of gland illustrating the possible process in cannabinoid localization in secretory cavity. A phenol glucoside is transported into a disc cell and stored as free phenol in the vacuole. Terpene is synthesized by the specialized plastid, lipoplast, in disc cells. These precursors, terpene and phenol, react to form cannabinoids at the plasma membrane surface or in the wall whereupon they appear in the secretory cavity.
As described in the cannabinoid pathway, these dimeric compounds consist of terpene and phenolic components. The abundant secretory activity of the disc cell plastids, and knowledge that this organelle does synthesize terpenes, suggests that they contribute the terpene component. Our detection, in previous studies, of abundant phenol in whole glands, and knowledge that phenols accumulate in vacuoles of cells, suggests that this cell feature may contribute the phenol component. Phenols are transported in the plant as glycosides and, when becoming localized in a cell vacuole, they accumulate there upon dissociation of the sugar moiety which returns to the cell cytoplasm.
We hypothesize that terpenes and phenols, when released from their respective sources, accumulate at the plasma membrane and cell wall interphase where enzymes dimerize these compounds into cannabinoids. It is necessary to determine enzymes involved in cannabinoid synthesis. Such an enzyme, when available, can be prepared as an antibody probe that can be used to identify more precisely the locus of cannabinoid, and THC, synthesis. Glands represent unique structures, and can be utilized to broaden our understanding of cannabinoid synthesis and aid in our effort to reduce the cannabinoid content of Cannabis strains for production of industrial hemp.
CONCLUSIONS
1. THC accumulated in abundance in the secretory cavity where it was associated with the: a) cell walls, b) surface feature of secretory vesicles, c) fibrillar material released from disc cell wall, and d) cuticle. It was not associated with the content of the secretory vesicles. The association of THC with structural components, particularly the wall, fibrillar matrix and surface feature of vesicles, suggests that it may be chemically bound to them rather than being free in the cavity. If THC and other cannabinoids are bound to components in the cavity, their presence and movement may require a source of energy in the cavity. Additional studies are necessary to determine their bound or free status.
2. Little or no THC was detected in the cytoplasm of the disc cells. This suggests that the terpene and phenol precursors, which must occur in the disc cells at some interval, may form the cannabinoids at the surface of the plasma membrane, or in the cell wall facing the secretory cavity.
3. Some THC was present in the cell walls of other cells. Genes for cannabinoid synthesis are present in all cells of the plant, but tissues other than glands produce low levels of these compounds.
4. Reduction or elimination of glands by mutation procedures will reduce significantly the quantity of THC in the plant. However, the pathway for cannabinoid synthesis is controlled genetically: glands are specialized to synthesize high levels of cannabinoids. Thus, a glandless plant can be expected to synthesize very low levels of cannabinoids. We do not know the roles of cannabinoids in the glands. They may be involved in some way with "protection", or other role. The absence of glands may or may not alter the functional role. Since other cells also synthesize these compounds, at very low levels, the quantity may be sufficient to perform the functional role. Therefore, a glandless mutant(s) would serve our purpose to reduce the THC concentration for utilization of such strains in the hemp industry.
ACKNOWLEDGEMENTS
This study was supported with stipends from the Korean Ministry of Education through Research Funds, 97-BSRI-4441 (ESK) and the Indiana University Faculty Research Program (PGM). We appreciate the generous gift of THC antibody provided by Roche Diagnostic Systems, NJ. Drug Enforcement Administration registration number PI00433113 (PGM).
REFERENCES
Hammond, C. T. and P. G. Mahlberg. 1973. Morphology of glandular hairs of Cannabis sativa L. from scanning electron microscopy. Amer. J. Bot. 60:524-528.
Hammond, C. T. and P. G. Mahlberg. 1977. Morphogenesis of capitate glandular hairs of Cannabis sativa L. (Cannabaceae). Amer. J. Bot. 64:1023-1031.
Hammond, C. T. and P. G. Mahlberg. 1978. Ultrastructural development of capitate glandular hairs of Cannabis sativa L. (Cannabaceae). Amer. J. Bot. 65:140-151.
Hammond, C. T. and P. G. Mahlberg. 1990. Thin-layer chromatographic identification of phenol in the glandular secretory system of Cannabis sativa L. (Cannabaceae). Ind. Acad. Sci. 98:201-209.
Hammond, C. T. and P. G. Mahlberg. 1994. Phloroglucinol glucoside as a natural constituent of Cannabis sativa. Phytochemistry 37:755-756.
Hemphill, J., J. Turner and P. G. Mahlberg. 1980. Cannabinoid content of individual plant organs from different geographical strains of Cannabis sativa L. (Cannabaceae). Jour. Nat. Prod. 43:112-122.
Kim, E. S. and P. G. Mahlberg. 1991. Secretory cavity development of glandular trichome of Cannabis sativa L. (Cannabaceae). Amer. J. Bot. 78:142-151.
Kim, E. S. and P. G. Mahlberg. 1995. Glandular cuticle formation in Cannabis (Cannabaceae). Amer. J. Bot. 82: 1207-1214.
Kim, E. S. and P. G. Mahlberg. 1997a. Immunochemical localization of tetrahydro- cannabinol (THC) in cryofixed glandular trichomes of Cannabis (Cannabaceae). Amer. J. Bot. 83: 336-342.
Kim, E. S. and P. G. Mahlberg. 1997b. Plastid development in glandular trichomes of Cannabis (Cannabaceae). Mol. and Cells 7: 352-359.
Lanyon, V., J. Turner and P. G. Mahlberg. 1981. Quantitative analysis of cannabinoids in the secretory product from capitate-stalked glands of Cannabis sativa L. (Cannabaceae). Bot. Gaz. 142:316-319.
Mahlberg, P. G., J. Turner, J. Hemphill and C. Hammond. 1984. Ultrastructure, development and composition of glandular trichomes of Cannabis, pp. 23-51. In: Biology and Chemistry of Plant Trichomes, E. Rodriguez, P. Healey, and I. Mehta (eds.), Pergamon Press, NY. Mahlberg, P. G. and E. S. Kim. 1991. Cuticle development on glandular trichomes of Cannabis L. (Cannabaceae). Amer. J. Bot. 78:1113-1122.
Mahlberg, P. G. and E. S. Kim. 1992. Secretory vesicle formation in glandular trichomes of Cannabis sativa L. (Cannabaceae). Amer. J. Bot. 79:166-173.
Turner, J., J. Hemphill and P. G. Mahlberg. 1977. Gland distribution and cannabinoid content in clones of Cannabis sativa. Amer. J. Bot. 64:687-693.
Turner, J., J. Hemphill and P. G. Mahlberg. 1978. Quantitative determination of cannabinoids present in individual glandular trichomes of Cannabis sativa L. (Cannabaceae). Amer. J. Bot. 65:1103-1106.
Turner, J., J. Hemphill and P. G. Mahlberg. 1980. Trichomes and cannabinoid content in developing leaves and bracts of Cannabis sativa L. (Cannabaceae). Amer.J. Bot. 67:1397-1406.
Turner, J. and P. G. Mahlberg. 1984. Effects of sample treatment on chromatographic analysis of cannabinoids in Cannabis sativa L. (Cannabaceae). J. Chromatogr. 283:165-171.
To go to the Table of Contents, click here.
To go to Part 1, Canada, click here.
To go to Part 2, Fibre & Food, click here.
To go to Part 3, International, click here.
To go back to Part 4, Research, click here.